|
|
|
ABSTRACT
Declining numbers of oceanic zooplankton cannot be entirely accounted for by the climate-warming-induced slacking of upwelling
currents. For example, the North Atlantic ocean offers a picture today of unusually low zooplankton numbers in the presence of
unusually high phytoplankton numbers, a finding which runs directly contrary to expectations. The life histories of common zooplankton
organisms (e.g. copepods) reveal that while juvenile stages are herbivorous, adult stages are not. They are frequently omnivores
or carnivores. The reproduction of herbivorous zooplankton therefore is not solely related to the availability of plant food.
The abundance and quality of carnivorous food available to the adult stages also impacts directly on the successful reproduction
of the herbivores.
Pelagic spawn released by marine fish and invertebrates is accessible food to these carnivorous zooplankton. The massive reduction
in "spawning stocks" of multiple marine species that has been induced by human fishing is therefore plausibly implicated in the
current declining numbers of zooplankton. Spawn production in the ocean has obviously declined with the elimination of spawning
fish, and this gradual withdrawl of a rich carnivorous food supply to the zooplankton may have negatively affected their ability
to reproduce. It also follows that total new organic production (and carbon fixation) in the marine system will have been reduced.
The widespread declining trend in age and size at maturity in marine fish and invertebrate species possibly represents a systemic
reaction to the falling density of zooplankton, in the form of an increased effort to produce spawn in compensation.
If fishing has "undercut" the zooplankton in this manner, and it seems entirely plausible that it has, then one "unintended,
unanticipated negative consequence" of continued fish removals will be the collapse of virtually the entire marine ecosystem.
Continuing to focus our explanatory efforts solely on temperature changes and "global warming," despite remarkable inconsistencies
between predicted and observed effects, merely continues a long-ingrained human thought pattern of denial...denial of the full
impact of our ruthless destruction of marine life...and perhaps denial that many of our earlier theories about how the ecosystem
functions are now proving to be, to a rather significant extent, incorrect. For example, the concept of energy flowing in only
a single direction in marine systems, upwards through successively higher trophic levels, is challenged by consideration of the
significant quantity of energy that is actually channelled directly downwards in the form of marine spawn. Although it inevitably
dissipates, energy can be seen to follow many circular routes within the ecosystem, cycling along with nutrients. In this way,
animate marine life in total acts as a catalyst for the continuation of high rates or primary production...a greater abundance
fish in the sea, therefore, ultimately results in a higher rate of marine carbon fixation.
Oceanic zooplankton is in serious decline. While the general public may not yet raise an eyebrow at this news, marine scientists
recognize it as an ominous sign. The decline of zooplankton is an unpredicted and ominous development in marine ecology. Estimates
of the abundance of zooplankton have only been done relatively recently, with the longest scientific data sets including perhaps
50 years of measurements. And the declining trend seems to span most of that period. Zooplankton abundance before 50 years ago,
and before the start of commercial fishing, is unknown. Following the publication of a research article in 1995 (Roemmich and
McGowan, Science, Vol 267, pp 1324-1326) the discovery was widely reported that "Since 1951, the biomass (living weight)
of zooplankton in the waters off the coast of southern California has decreased by 70%." (www.grico.org). Indications of steady,
long term declines in zooplankton have also been found in other ocean areas. A study of the baleen of bowhead whales in the Bering
Sea recently revealed a 40 year declining trend in the nutritional intake by these plankton feeders. (D. Schell’s research, reported
at www.discovery.com) Substantial zooplankton declines have also been recorded in the Baltic Sea and in the North Sea. (IGBP/GLOBEC)
The most recent report is possibly the most worrying. A plankton survey of the North Atlantic was carried out by a team of British
scientists during November-December of 2001. Their preliminary results:
"The results have been worrying. It was anticipated that levels would be around 50,000 per square metre in a column of water.
Instead, between 5,000 and 10,000 specimens were recorded. ‘That is an order-of-magnitude difference, and indicates something
very serious may have occurred,’ said Dr. Phil Williamson (of East Anglia University) … ‘This is deeply worrying. We don’t
know why zooplankton numbers have plummeted, although global warming looks like the best candidate. What is certain is that
removing the bottom link from the ocean’s food chain could have profound and unpleasant results,’ he told The Observer."
(from an online news item, Dec. 4, 2001, posted at www.fis.com )
The most favored explanation for the decline of zooplankton has been that is it an effect of "global warming." Regarding the
study off the coast of California, the 70% decline in abundance since 1951:
"During that same period, the surface layer of the ocean warmed, by more than 1.5 degrees C in some places, and the temperature
difference between the layers increased. This increased stratification resulted in less lifting of deeper waters by wind-driven
upwelling. A shallower source of upwelled waters provide less inorganic nutrients for new biological production and hence supported
a smaller zooplankton population." (www.grico.org)
This explanation for the lower numbers of zooplankton is initially a plausible one. The sequence of events is believed to be
that weaker upwelling forces make less nutrients available for the stimulation of phytoplankton growth. The effect of this on
the herbivorous zooplankton is that they therefore experience a degree of food shortage, and the result is a decline in their
numbers. Since the mechanism of zooplankton decline is believed to be a shortage of phytoplankton food, presumably the Californian
study also found relatively low levels of phytoplankton. But I have not found reference to this particular aspect of the story.
One finding that would seem to strongly contradict their hypothesis, however, would be if an increase were discovered in the
standing stock of phytoplankton.
Declining zooplankton populations in the presence of normal, or higher than normal, levels of phytoplankton growth, would
imply that some other factor is involved and also exerting a negative effect on the growth of zooplankton.
This scenario is exactly the one that has been discovered by Canadian scientists monitoring the ocean off Atlantic Canada,
and this finding complicates the larger picture. Another explanation is needed. The Canadian Department of Fisheries and Oceans
has plankton monitoring data sets for the Scotian Shelf that extend back to 1961. Trends were summarized in a recent report (Stock
Status Report G3-02
(2000)):
"- The colour index for the Scotian Shelf derived from the Continuous Plankton Recorder (CPR) data showed a large increase in
the 1990s compared to 1961 - 1975 that was due to an increase in the numbers of diatom and dinoflagellate phytoplankton. - The
CPR numbers of the early stages of the copepod C. finmarchicus and the total copepod numbers showed a general downward trend
in the 1990s compared to 1961 - 1975. - The CPR total numbers of krill (euphausiids) in the 1990s were below the long-term mean."
So phytoplankton are up, while zooplankton are down; this is the unexpected trend that has emerged over the last decade…what
explanation is offered? In this case, with zooplankton declining in the ocean off Atlantic Canada as it is elsewhere, there is
no suggestion that the phenomenon has been triggered by "warmer surface waters/decreased upwelling and decreased lifting of dissolved
nutrients from deep waters," because there is nothing to indicate that physical mixing of water layers has decreased in this
area. Instead it appears, by the occurrence of extensive blooms of phytoplankton, as if adequate amounts of bottom water and
nutrients are still being physically raised to the surface. The DFO report offers a reference to a transient period of colder
water, but no convincing explanation:
"The years 1991 to 1998 were a period of increased phytoplankton levels and reduced population levels of copepods and krill
on the Scotian Shelf. The earlier 1990s were years of cold water temperatures on the northeastern Scotian Shelf. These cold temperatures
are believed to have played a role in the changes seen in the phytoplankton and zooplankton communities." (DFO, 2000)
One problem with this explanation is that the cold temperatures noted in the early 1990s were over by the mid 1990s, and water
temperatures in recent years have been at or slightly above long term means. The same cold temperatures in the early 1990s were
also once "believed to have played a role" in the crash and failure to recover of the once huge groundfish stocks in the same
area. It was predicted in the early 1990s that cod and other fish would begin to grow better once the temperature returned to
normal…but that is not what happened. The fish stocks overall have simply continued to decline.
It seems more likely that there may exist a direct link between the disappearing fish stocks and the declining zooplankton
stocks, than that a transient cold spell a decade ago was an important cause of either.
One side of the feeding relationship between fish and zooplankton is very well understood, and this is the fact that fish
eat zooplankton. Virtually all fish depend on zooplankton as a food source during their early life stages, and some fish
feed on it for their whole lives. But a close look at the habits of zooplankton reveals that, possibly to a very significant
extent, many zooplankton also eat fish. By ‘eating fish’ I mean consuming their offspring. Zooplankton are significant
predators on the eggs and larvae of fish species that release pelagic eggs, and this includes the majority of fish species. It
is important to note that many species of zooplankton (including the highly abundant calanoid copepods and chaetognaths) commonly
shift their diet from herbivorous as juveniles to omnivorous or carnivorous diets as reproductive adults. And that the numbers
of their offspring is known to be directly related to their level of nutrition. Better fed copepods produce larger batches of
eggs. (Steidinger and Walker, 1984) Therefore, the successful reproduction of these ‘herbivorous’ zooplankton depends not only
on an adequate supply of phytoplankton, but also on an sufficient supply of appropriately sized ‘animal’ food. Phytoplankton
functions best, it seems, only as ‘baby food’ for many zooplankton species, therefore a vigorous population growth cannot be
expected if that is the only food that is provided in abundance.
The long recognized spring bloom of phytoplankton in the North Atlantic is well known to be followed shortly thereafter by a
marked increase in the zooplankton abundance…therefore simple cause and effect have been assumed. While it is fairly certain
that the heightened supply of phytoplankton contributes to the zooplankton population growth, it bears pointing out that a supply
of planktonic carnivorous food also normally ‘blooms’ in the spring, coinciding with the blooms of phytoplankton and zooplankton.
The release of pelagic spawn by a multitude of marine species, both fish and invertebrates, also increases sharply at this time
of year. Most of this spawn is consumed within the plankton. While zooplankton certainly consume many carnivorous foods besides
spawn (e.g. smaller zooplankton, bacteria), fish eggs may represent the most nutrient dense items available, and the infusion
of billions of floating eggs and sperm into the plankton may represent a significant source of food to these tiny carnivores.
The combination of ‘physical forcing’ (via the mixing induced phytoplankton bloom) and ’biological forcing’ (via the seasonal
‘bloom’ of floating spawn) may therefore be what is needed to produce optimal growth of the zooplankton population.
The carnivorous food requirements of common, abundant zooplankton species is a subject that has received relatively scant scientific
attention:
"We have insufficient knowledge concerning the impact of copepod carnivory in the marine ecosystem. Emphasis in quantitative
studies has been almost entirely on herbivorous feeding, partly because of the outmoded concept that all copepods are primarily
herbivorous and partly because quantitative estimates of carnivory are much more difficult to undertake." (Charles C. Davis,
in Steidinger and Walker, 1984)
Dramatic declines in spawning stocks have occurred in virtually all commercially fished species in Atlantic Canada (and elsewhere).
It follows that the quantity of their spawn released into the ocean has declined in a similar manner. Disturbingly, unexploited
spawn producing species have also recently been noted to be in decline, for example look at
barnacles.
Could this downturn in overall pelagic spawn production be large enough to be felt as a food loss by the zooplankton community?
"PHYSICAL FORCING" vs "TIGHT CYCLING OF NUTRIENTS" - two different dynamics known to drive marine primary productivity….is
it one or the other, or is it both?
Two processes are described in marine ecology texts as "driving" marine primary productivity. The first, mentioned already,
is "physical forcing." In areas of the world ocean where periodic mixing of deep, nutrient rich bottom water with surface waters
occurs, this physical mechanism has been recognized as triggering seasonal bursts of phytoplankton growth or "primary productivity,"
and this has then been viewed as the major determinant or ‘driver’ of productivity in those areas. The perception has been one
of biological productivity being forced by a purely physical process. This effect is much more greatly pronounced in some areas,
including the "upwelling" areas and the seasonally mixed temperate seas, such as much of the North Atlantic. Scientists have
determined that the deep waters of the world ocean contain a truly vast reservoir of dissolved inorganic nutrients; quantities
far in excess of the total nutrient inventory held in the photic zone exist below the zone where photosynthesis is possible.
Only a very small fraction of this reservoir is raised to the surface annually by mixing, and organic matter sinks continuously
back down to the depths, so the content of this reservoir is thought to be basically a constant. Given stable climatic conditions,
where forces causing physical mixing of waters are a consistent phenomenon, it has been believed that these conditions will ensure
a generally constant rate of marine primary productivity, since approximately the same amount of nutrients will be raised to
the surface by mixing each year…and the quantity of new organic life formed is believed to be directly related to the quantity
of nutrients raised. This underlies our beliefs that ‘annual marine new productivity’ is a constant, and that, while species
shifts may be induced by fishing, nutrient depletion of the entire system by fish extraction is impossible (since the bulk of
nutrients removed by fishing represents only a very tiny fraction of the nutrients contained within the ocean as a whole.)
The understanding of the physical forcing dynamic has probably also contributed to the (mistaken) general belief by fishery
mangers that the sea is capable of producing food for us endlessly. (The idea is that the top levels of the food web can be removed
by fishing continually without causing any weakening or deficit in the system overall. The worst result foreseen from "over fishing"
has been that the fished species will disappear and that the (steady) organic production of the ocean will then simply be channelled
into species that we do not care to eat (such as plankton, microbes, and jellyfish)…not that the total capacity for new organic
production might be hampered. Since "physical forcing" could be counted on to always provide the nutrients needed to build new
fish, no need to fertilize the sea for this reason has been appreciated by humans. Fishing, unlike all our terrestrial food harvesting
endeavours, appeared to us to be capable of being sustained indefinitely…just continual harvesting with no requirement for nutrient
replacement or fertilization…certainly a terrestrial equivalent to this has never been found…and now it appears that this view
of the food production potential of the ocean was just too good to be true...)
It seems more likely now that the total quantity of new organic material that can potentially be formed in the photic zone depends
not only on the amount of nutrients regenerated by physical mixing of the deep waters, but also on the total quantity of living
organic material (fish, for example) that already exists within the photic zone. The living assemblage itself effectively acts
as a catalyst for new production. Energy and nutrients held within adult organisms are regularly channelled 'downward' in a manner
that helps to stimulate growth and reproduction of zooplankton and phytoplankton, and accelerate uptake of new energy into the
system...spawn production and consumption is an unrecognized route by which this dynamic occurs. The more organisms that inhabit
the sea, the greater the capacity of the ecosystem as a whole to absorb whatever dissolved nutrients are made available, and
to maximize the resulting potential for new organic production.
The discovery of highly productive marine ecosystems (coral reefs) in the shallow, nutrient-poor tropical waters initially seemed
to present a conundrum. Because physical mixing generally does not occur in these areas, it was not initially obvious to scientists
what process was driving such high productivity. Study of the tropical organisms, however, revealed that the secret to their
productivity lies in the complexity of the elaborate symbiotic relationships that exist between species. ‘Give and take’ relationships
exist on many levels, with the effect that the living community as a whole has a strong tendency to conserve nutrients once it
gains possession of them. Nothing is wasted; nutrients are cycled repeatedly, allowing ongoing photosynthesis in the absence
of physical mixing, and boosting productivity to higher levels. This pattern has been described as "tight recycling of nutrients"
and it allows for the continuation of significant levels of primary production in very thermally stratified, nutrient-poor tropical
water. In contrast to most scientific writing on temperate fisheries, standing stocks of fish have actually been perceived as
being significant nutrient reservoirs in scientific analyses of tropical fisheries. Their removal has been described as being
a possible cause of lowered productivity due to their role in nutrient cycling. (Polunin and Roberts, 1996).
But tropical and temperate marine species belong to the same families of organisms…all are simply variations on a few common
themes…as are the varieties of marine ecosystems on the planet. And if tropical marine assemblages tend to conserve and recycle
nutrients as an organizing principle that augments (or completely replaces) physical forcing of productivity, why would this
not also be true of their temperate cousins? The coral-zooxanthellae symbiosis is an arrangement unique to the tropics, but what
it accomplishes - an efficient short circuit that neatly passes vital nutrients back and forth between plant and animal - is
not.
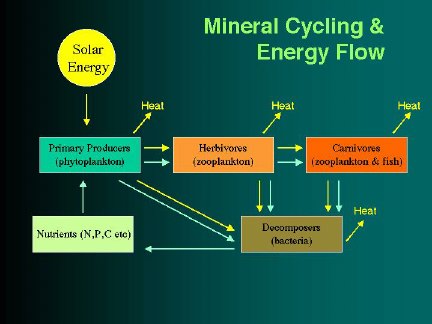
Food web diagrams for marine ecosystems typically have emphasized the movement of fixed carbon (or energy) up through successively
higher trophic levels, as smaller organisms are consumed by larger ones. (Very often these diagrams show the pathway leading
to commercial fishery production…and coming to an end there.) Although a greater complexity is now generally acknowledged, at
least to the extent that fish species are recognized as feeding at different trophic levels at different stages of their lives,
what is notably absent from marine food web diagrams are the short cycles that occur within the web, short feedback loops involving
direct transfer of nutrients and energy "down" from higher trophic levels to levels below. Many such cycles can be appreciated
once the consumption of spawn is factored in. (This further confuses the "trophic level" concept, since eggs produced by large
fish are food that has passed through several ascending levels leading to the parent fish, but when they are consumed by zooplankton…at
which "level" is the zooplankton actually feeding?)
Studies of egg and larval mortality reveal the extent of their consumption by members of the zooplankton:
"The high rates of mortality of eggs and early larval stages (as high as 95% per day, but more typically 30 to 40% per day)
suggest that predation must be intense…Pelagic invertebrate predators such as ctenophores, chaetognaths, medusae, siphomedusae,
euphausids, copepods, amphipods, and dinoflagellates have all been reported to consume fish eggs and early larvae….Information
obtained from laboratory studies indicate that euphausids and calanoid copepods are potentially serious predators of fish eggs
and larvae…Some hyperid amphipods have been implicated as serious predators on fish eggs and early larvae." (Steidinger and Walker,
1984, p 130)
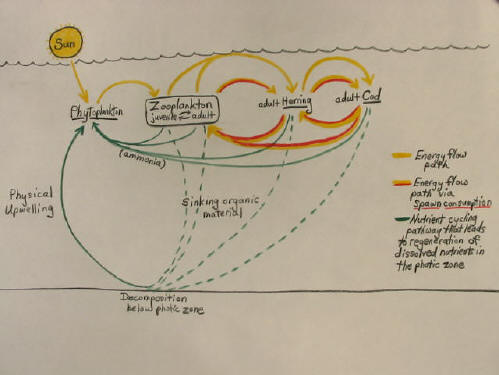
What do the "tight cycles" look like?
One "tight cycle" involves the direct fertilization of the phytoplankton by all other creatures in the sea. Marine organisms
living at all trophic levels above the primary producers excrete dissolved nutrients into the water that are ideally suited to
fertilizing the phytoplankton. Very often this is ammonia, which all fish excrete from their gills. This has sometimes been recognized
in the literature as a significant route of nutrient regeneration in surface water, but usually only the contribution of the
zooplankton has been acknowledged. Recently a "microbial loop" dynamic has been appreciated in marine food webs, one effect of
which is the regeneration of nutrients into the photic zone. (Nybakken, 1997) All larger fish, however, also contribute in the
same manner to this ongoing stimulation of primary production.
"The excretions of the zooplankton also contain nitrogen, phosphorus and other substances which can be used by phytoplankton
and it has only recently been realized how important a part they may play even in surface waters. In Long Island Sound it was
thought that during active phytoplankton growth in summer half the nitrogen used came from zooplankton excretion. An advantage
to the plants is that this is a quick process; bacterial decay is much slower." (Orr and Marshall, 1969, p 47)
The classic nutrient return route of ‘sinking/decomposition/upwelling’ is often included in food web diagrams to show the recycling
of nutrients excreted by fish, while the shorter feedback loops, such as in the preceding example, circuits capable of significantly
amplifying the total rate of production, have generally not been included. (The ‘sinking’ route is commonly followed by some
fish excretions, notably feces and detritus, but other excretions, ammonia and spawn, have a strong tendency to stay within the
photic zone.)
Patterns often repeat in nature. The dual and complementary effects that zooplankton have on phytoplankton (predation and fertilization)
are repeated in turn in the effects that their consumers have on the zooplankton population. Of the many fish and invertebrate
species that feed on zooplankton, the vast majority also release pelagic spawn, and their eggs and larval stages then serve to
offer high quality food in portions small enough to be captured by many of the larger zooplankton organisms, This short feedback
loop, a level removed from the primary producers, has the potential to increase the efficiency with which the web as a whole
incorporates the products of primary production into the carnivorous web. This is because well nourished (carnivorous) adult
zooplankton will produce larger numbers of herbivorous offspring, thereby grazing the phytoplankton more completely and allowing
less of the products of photosynthesis to sink to the bottom. Thus the existence of the ‘downside’ of the second loop (fish spawn
nourishing zooplankton) enhances the overall efficiency of the first loop (zooplankton consuming and fertilizing phytoplankton).
Therefore, the existence of healthy, intact populations of consumers higher in the food web can be seen to enhance overall productivity
- such an ecosystem is better positioned to take advantage of nutrient pulses provided by physical forcing and enhance and prolong
the resultant productivity by "tight recycling of nutrients" or "biological forcing."
The ecological significance of marine spawn deserves a closer look.
Most scientific attention that has been paid to fish eggs and larvae has been focused on the survivors, often on the tiny fraction
that ultimately "recruits" to a commercially exploited fish stock. But "recruitment" is the rare outcome, practically all spawn
produced effectively serves a different ecological purpose. By becoming part of the zooplankton, it nourishes either other zooplankton
organisms or the species that feed on zooplankton, such as juvenile (or adult) fish.
Once the consumption of spawn and juveniles is added to the marine food web diagram many "tight" nutrient cycles can be appreciated
inside the overall scheme, and all of these short feedback loops have the ‘side-effects’ of conserving nutrients within the living
web, while steadily regenerating nutrients directly into the water of the photic zone. A lot of trading back and forth occurs
between various species. The labels "predator" and "prey" can be seen to alternate between species, depending on which life stage
is being considered.
Numerous examples of ‘give and take’ circuits can be cited, feeding relationships often clearly swinging both ways. As has been
already described, fish species that feed on the common zooplankton such as copepods, for instance, are the same fish that ‘lose’
eggs to copepod predation...but symbiotic feeding relationships may look still different from that. Seemingly independent species,
when viewed only as adults, can sometimes be seen to engage in reciprocal feeding relationships. Barnacles and fish initially
appear to have no interdependency in that neither one consumes adult members of the other group. A look at the pattern of spawn
consumption, however, reveals that the juvenile, pelagic life stages of the barnacle are commonly consumed by plankton-feeding
fish…and also that fish eggs and larvae are of a size that can be readily captured and consumed by adult barnacles. So…do fish
eat barnacles?…and do barnacles eat fish? Could a severe reduction in the spawning stock biomass of fish result in a missing
food source that could have a negative impact on barnacles? (Barnacles,
incidentally, are also showing marked declines in some areas.)
In temperate seas groundfish (gadoids, flatfishes) and small schooling pelagics known as "baitfish" (herrings, capelin, etc.)
are two historically very highly abundant fish families. A large fraction of the total ‘fish production’ in the ocean was therefore
channelled through these two species groups. A short cycle by which nutrients pass back and forth between these two groups appears
to have been included in the design of the system. The "baitfish" (herrings) are primarily plankton feeding fish as adults, that
are ideally equipped to strain out free floating fish eggs from positions high in the water column. The eggs of the groundfish
(cod, haddock, etc.,) are typically pelagic, floating high in the water column where they present themselves as an ideal food
source for the "baitfish."
"The predators having the greatest impact on (pelagic) fish eggs and larvae are probably schooling (pelagic) juvenile and adult
fish…Much of this predation is on newly spawned eggs as they are released and still highly concentrated. While these predators
are general filter feeders, they seem to be attracted to spawning sites specifically for the purpose of feeding…Planktivorous
fish are known to be important predators of fish eggs and larvae, and a large school may well eliminate a patch of fish eggs
or larvae…"(Steidinger and Walker, 1984, p 131)
So the bottom fish send their eggs to the surface to be largely consumed by the pelagic fish. But there is a neat reversal of
fate when the herring and capelin spawn. In what appears to possibly be a reciprocal ‘pay back’ design, the eggs of these fish
sink to the bottom where they adhere to surfaces and to one another. The eggs of herring are piled many layers deep, to the point
of suffocating those buried most deeply. This ‘reproductive strategy’ (producing a large mass of demersal eggs) on the part of
the herrings may or may not confer particular advantages on their newly hatched young, but it clearly does confer an advantage
on the groundfish that normally feed on them. Groundfish are unable to filter planktonic particles, so they can derive no benefit
from free floating pelagic eggs, but they can and do take advantage of the clumped together sunken herring eggs. Groundfish are
also well known to "be attracted to spawning sites specifically for the purpose of feeding." Dragger fishermen, at least in some
inshore areas in Atlantic Canada, are well aware of the tendency of groundfish to congregate at herring spawning sites. A bottom
trawl hauled through such a site will typically come up coated in sticky herring spawn and full of "spawny" ground fish (their
bellies filled with herring eggs). The fishermen call this a "spawn tow." Since they cannot effectively prey on single eggs,
groundfish are unlikely to "eliminate" patches of herring eggs. Their feeding may ultimately improve the survival odds of the
young herring, however, since their feeding will break up the thick clumps and allow the survivors to be better oxygenated than
they would have been in the undisturbed egg mass. The ‘reason why’ pelagic fish produce demersal eggs has been difficult to say
with certainty. Demersal eggs experience lower predation mortality than do pelagic eggs, so apparently the groundfish are unable
to consume the baitfish spawn to the same extent as the herring can polish off the floating cod spawn. Their overall relationship
is not really unbalanced in favor of the herring however, since the feeding balance tips the other way when mature cod feed on
older juvenile and adult herring. In the final balance, it seems that both families of fish evolved strategies to practice "sustainable
harvesting" of the other…considering their successful coexistence for millions of years, and the immense numbers of these fish
that existed in the sea prior to the onset of human exploitation.
These few examples offer glimpses of ‘tight recycling of nutrients’ as it is done among the temperate zone fish stocks, and
also clues as to the reason why so many fish stocks area failing together. They evolved together and none was meant to, nor is
it capable of standing alone.
Our first impression of some fish behaviours, such as the piling up of thick layers of herring eggs, might be that, in some
aspects, their approach serves to defeat their purpose (which we assume to be simply maximizing the survival rate of their young)…but
in fact these fish are doing things very efficiently, as they have done for millions of years…the problem is that we may not
yet have accurately identified all of their ‘purposes.’
HOW SIGNIFICANT MIGHT FISH SPAWNING BEHAVIOR BE AS A NUTRIENT CYCLING STRATEGY IN THE OCEAN?
This is an important question. Does the spawn of fishes represent a minor and relatively insignificant fraction of the zooplankton…or
is the biomass and nutrient transfer involved large enough to be considered significant?
How important is a food that is only seasonally available? Can it represent a important portion of the diet, or is fish spawn
only a trivial and occasional meal, not enough to have an effect on the overall nutritional health of the consumers? Considering
that most food supplies in nature, certainly those in the higher latitudes at least, are only made available to their consumers
on a seasonal basis, the fact that spawn only appears ‘in season’ does not argue against its importance as a food.
Are the quantities of food involved in the consumption of fish eggs and larvae significant? (Bearing in mind that all we have
to assess at this time is a severely altered and degraded marine ecosystem, it’s hard to say with any certainty what role fish
eggs might have originally played in the diet of zooplankton, or what the concentration of fish eggs in seawater might have been
in the unexploited system…regardless however of the actual ‘concentration’ of eggs that might have accumulated in the water,
the more significant parameter is the ‘supply’ of eggs, which obviously was much greater before the destruction of the spawning
stocks.)
Older reports can provide glimpses into the quantities of fish eggs that were once released into the sea:
"The occurrence of spring herring spawning beds in shallow waters of the littoral zone often results in eggs being washed ashore
during gales. Prince (1907) stated that, in some years, herring eggs were heaped knee high for kilometres along the New
Brunswick shore after severe gales. Escuminac-area fishermen reported that enough eggs were washed ashore to be collected for
fertilizer as recently as the early 1960s. The negligible amounts of beached material during our surveys from 1980 to 1984
most likely reflected the decline in herring stocks and the small amount of spawn compared to recent years." (Messieh, 1988)
A significant fraction of marine zooplankton apparently still does consist of the eggs and larvae of creatures who are only
there temporarily as juveniles. Discussions on this topic often group together the larval stages of fish and invertebrates due
to their similarities; for example:
"Meroplankton. Only a part of life (usually during larval stage) is spent as plankton. After metamorphosis, it changes to benthic
or nektonic form. Due to its great abundance, this group of plankton (mostly larvae) occupies an important place in the plankton
community, especially during the breeding season. Since they appear periodically at definite seasons (spring, summer or autumn),
they are called seasonal plankton." (Zhong et al. 1989, p 4)
Therefore, it appears that a severe decline in the numbers of mature fish and invertebrates in the sea might result in a noticeable
lack of an "important" segment of the zooplankton. Seasonal availability covers most of the year. Therefore, if the egg or larval
consumer is indiscriminate as to species consumed (which they are well known to be), the overall opportunity to consume this
type of food is fairly substantial.
How much spawn do fish produce?
Of course this varies with the species , but it appears to range between approximately 10% and 30% of the biomass (wet weight)
of the fish. The percentage is also referred to as the "gonadosomatic index" (GSI). 20% may be a reasonably representative figure.
Considering that 20% by weight of the average spawning fish stock is diverted to the zooplankton, the bulk of the nutrient transfer
might start to look significant…especially if we consider the immense spawning stocks of fish that existed prior to human exploitation.
For every 100 tons of living "spawning stock biomass" (SSB), approximately 20 tons will be annually converted to zooplankton.
The fish that divert the highest proportion of their resources to this use are the biggest, oldest spawners. This is well known.
The larger sized fish put less energy into somatic growth and divert a larger fraction of their intake into ‘reproduction.’ Their
own bodies growing only relatively slowly once they reach the larger sizes, the biggest spawners achieve not only the highest
GSI and the highest number of eggs, but also the highest quality eggs and larvae (more yolk, stronger larvae) (Hempell). Both
the quantity and the quality of fish eggs in the sea was therefore once much higher.
(The real significance of the large spawners, including their ecological role as efficient replenishes of the zooplankton, appears
to have been lost on fishery managers. With a view only to maximizing the growth rate of the commercially desired fish flesh,
the standard short-sighted view has been that removing the "old, slow-growing component" of a fish stock actually enhances the
production of fish. Therefore, unfortunately, the largest spawners have often been deliberately targeted and promptly destroyed
by fisheries. A stated initial management goal for newly exploited fish stocks was often to cut the population down to 50% of
the size of the virgin stock.)
Beyond the 20% by weight that mature fish divert annually to the zooplankton, the significance of the nutrient contribution
increases once the protein and energy content of roe is considered. Fish eggs are very high in protein (making extremely nutritious
food, as is well appreciated by their human consumers). The food value of fish eggs (by weight) is much higher than that of the
fish that produces them. The density of protein and energy in the eggs is such that the fraction of the total nutrient content
of a mature fish that is regularly diverted to the zooplankton can safely be assumed to be well over 20% of the total reserves
held within the body of that fish. Therefore all mature fish, and to a greater degree as they become older, divert a significant
portion of the nutrients and energy consumed into a recycling route that gives a direct boost to the plankton productivity and
is almost guaranteed not to be lost to sinking/decomposition.
The imperative to divert significant protein and energy stores into ‘reproduction/zooplankton replenishment’ appears to be quite
intense in fish. Following spawning, fish are well known to typically be in their lowest condition (fat content) of the year.
Protein previously incorporated into their own flesh may be drawn upon and diverted into egg production. This is evident from
the "jellied" flesh that has been noted in several groundfish species post-spawning. Cod in Atlantic Canada have recently been
deemed unmarketable due to the "jellied" condition of their flesh. This change in texture is the direct result of extreme protein
loss in these fish.
The diversion of protein and energy into egg (and sperm) production is therefore clearly a high priority biological activity
for fish.
A general trend of declining ages and sizes at maturity has been noted in commercially exploited fish stocks. Roe is appearing
in smaller specimens than ever before. And the reason for this change has presented another mystery. Hypotheses have been put
forward, such as the idea that early age and size at maturity has resulted from "size-selective culling" by fishing gear. The
change has occurred in both lightly exploited and heavily exploited species, however, which makes fishing gear effects rather
unlikely as the primary cause. Since commercially exploited fish species are at generally low abundance now, the idea has arisen
that they are spawning earlier somehow in reaction to the currently low numbers of their own kind. Somehow they feel a more urgent
need to reproduce, in an attempt to repopulate their own deleted species. But marine species currently at relatively high abundance
are also now maturing and producing eggs at smaller sizes, making the ‘intense urge to repopulate ones own (scarce) species’
hypothesis unlikely. An example in Atlantic Canada is the lobster. Lobster is still abundant; commercial landings are at historic
highs. But size at maturity has clearly been dropping in lobsters in recent years. (This has been reported by DFO, the FRCC,
and also by personal communication from local lobster fishermen and my own personal observation.)
It is possible that the biological trigger for early egg production is related to the depleted state of the zooplankton. Participation
in the zooplankton, even if only temporary, is common to the life histories of the majority of marine species. If they do not
survive this passage none of these species can continue. The contribution by all of them of large numbers of ‘extra’ spawn to
the sea ensured an appropriate food density for all of their surviving young. The zooplankton can be seen as functioning somewhat
like a ‘group life insurance policy’ for all of its players. But there has been a mismatch between premiums paid and withdrawls
from the fund for many years now due to the disappearance of great numbers of the larger spawn producers. As a result, premiums
have recently been raised for everyone…hence we see spawn production by ever younger organisms of all the involved species… The
exact trigger for the early onset of maturity might be a biochemical signal from the plankton, or it could be an internal thing
as simple as a slowing of the somatic growth rate, which is another widespread trend in marine life today.
THE VARIETY OF SPAWNING BEHAVIORS IN MARINE FISH: does this represent a co-operative effort to maximize the survival of
all, or is each species acting solely ‘in its own interest?’…And why are fish eggs always so small?
"Species composition and abundance of larvae, as in adults, change with the seasons, and this is apparently related to the
breeding seasons. In spring, summer and autumn, when the breeding of animals is in full swing, the larvae are abundant both in
diversity and quantity, forming the predominant portion of the plankton community. However, the peak of abundance appears
to vary with different species." (Zhong et al, p 426)
The suppliers of fish eggs and larvae to the ocean spread their contributions throughout the year. Coexisting fish species in
temperate zones stagger their spawning times to quite a large extent. Winter is the least favored time, although winter spawning
does occur (e.g. sand lance, pollock). Within single species, especially the dominant ones such as cod, the season of spawning
may be spread over several months. Others, notably herring, have stock components that spawn in spring and others that spawn
in fall. Larger individuals within a stock begin to release spawn earlier, so when the large standing stocks of older fish existed
in the sea, presumably spawning seasons were protracted to a somewhat greater degree even then they are today, extending the
egg feeding season for their consumers.
Why do fish stagger their spawning periods? To minimize the food competition experienced by their young perhaps, but perhaps
also to provide year round benefit to the consumers of fish eggs and larvae, including the zooplankton? The second result will
happen as surely as the first. In nature, seasonal supplies of food tend to be nicely synchronized to the times when they are
most needed by the consumers. This is a characteristic of interdependent species that have evolved together, and can easily be
seen in both terrestrial and marine ecosystems.
In northern temperate zones, at least, spring and summer are very popular times for spawning, most species (fish and invertebrates)
releasing their eggs at this time. Does this provide the best possible feeding environment for the young fish, or is a second
ecological purpose also being served by this choice of timing? The North Atlantic ocean experiences a major bloom of phytoplankton
in the spring due to strong physical forcing. Zooplankton also increases in response to this, as well as in response to the ‘spawn
bloom’ that also occurs at that time of year. Spring therefore should be a very good time for a small fish to be born that must
feed itself, even temporarily, on plankton. As the summer progresses, thermal stratification of the water column occurs and the
standing stock of phytoplankton becomes markedly diminished. Surface waters become relatively depleted of dissolved nutrients
and phytoplankton productivity drops off during the summer months. Many fish do not release their spawn until summertime however…and
timing their births to the season of lower phytoplankton density might at first seem to be less than ideal. This might place
the young fish at a feeding disadvantage compared to the spring hatched species. But is there another ecological benefit derived
from a large release of fish spawn during the summertime? The appearance of fish eggs and larvae in surface waters at this time
helps to ward off nutrient depletion of surface waters, which helps to maximize primary production by the phytoplankton during
the time of greatest sunlight exposure. It is interesting that the summer spawned eggs are most commonly the floating type. Fish
spawn released during summer months helps to sustain the growth of the zooplankton, which are then more able to both feed on
and to fertilize the phytoplankton. Continuing to make fertilizer available to the plants throughout the summer ensures their
best growth, and this goal is well served by the summertime spawning habits of so many fish.
The timing and habitat selection of the spawning habits of one species can often be seen to offer timely food supplies to another.
Beyond the examples given of summertime zooplankton augmentation by fish spawn, and the ‘groundfish-baitfish’ spawn feeding reciprocity,
many other happy coincidences can be found. For another example, the few surviving summer spawned groundfish larvae will typically
settle to the bottom after several months of life in the plankton…arriving there in early winter. The young, newly settled bottom
feeder arrives just in time to feed on the sticky demersal eggs of the sand lance that are released at that time. ‘Why’ would
the sand lance choose a winter spawning season? …And ‘why’ choose to produce a demersal egg? Who knows?… but it seems just possible
that at least part of the reason might be the provision of high quality, easily accessible food to tiny, juvenile groundfish
exactly at the time that they need it.
The continuum of levels of parental care by fish runs the gamut from advanced development of juveniles within the mother’s body
(similar to levels of care provided by some terrestrial animals), to some brooding activity and avoidance of predation risk for
the young…to the opposite extreme: reproductive strategies that amount to serving up the spawn precisely to suit the feeding
needs of the predators. Virtually delivering the young into the mouths of their chief predators seems to be a strategy followed
by some of the more successful fish families in the sea (groundfish and baitfish). The continuum of care has been recognized
and described in the literature, but just how ‘careless’ fish can be with the lives of their young has not generally been described.
Some spawning strategies seem to be designed first and foremost for the convenience of the spawn predator(!) Yet, this appears
to be one facet of a highly successful community of aquatic organisms that has evolved over millions of years, with all of the
necessary checks and balances in place to ensure continued survival. Their common dependence on the zooplankton, the nursery
and first food for all of their young, dictates the importance of continually maintaining this crucial element of the system.
Perhaps this is the reason that producing such huge numbers of tiny floating eggs evolved as the major method of animal reproduction
in the sea.
Why do even the largest fish produce such tiny, vulnerable eggs?
The probability of being consumed at the earliest life stages is so high that, only by dint of their sheer numbers, do a tiny
fraction of the offspring of fish avoid this fate and survive to maturity. Life as a tiny planktonic fish egg or larvae is usually
quite short, the risk of predation mortality at this stage being far higher than that faced by juveniles that have reached larger
sizes and ‘escaped’ from the perilous life in the plankton. Fish have evolved to cope with this awesome risk faced by their young
by producing them in huge quantities…or at least that is our usual interpretation of the reason why so many ‘extra’ eggs are
formed.
A minority of fish have evolved with reproductive strategies that avoid the extreme hazards of early planktonic existence as
juveniles (mainly sharks and their relatives). This is accomplished by a greater investment being made into each of a few offspring.
The dominant type of fish in the sea however, the ‘bony fishes’ or ‘teleosts’ overwhelmingly have opted to produce sheer numbers
of tiny offspring rather than investing more heavily in a smaller number of larger, better provisioned offspring (e.g. providing
more yolk per egg). But the reason for this has presented a bit of a scientific puzzle.
Scientific analysis of the energetics of this common reproductive strategy used by fish (broadcasting tiny eggs that hatch planktivorous
larvae) do not show it as being the most advantageous choice…or at least not the one that will give the highest rate of return
(survival of young) for the investment of energy and nutrients. Dispersal of offspring is thought to be an advantage conferred
on sessile invertebrates that use the ‘broadcast’ reproduction method, but for mobile adults such as fish, this cannot be argued
as an important advantage. It appears (to us) to be especially illogical for the largest fish to continue to produce such tiny,
vulnerable eggs and larvae. The giant bluefin tuna, for instance, a top predator (which grows up to 2000 pounds and 14 feet in
length), produces an egg that is the same size, or a tiny bit smaller than, that produced by tiny fish such as capelin and smelt,
small "baitfish" that grow to a maximum size of about 8 inches long. The eggs of all of these species are in a common size range
for many fish eggs, (smelt eggs are .05" in diameter, capelin eggs .04", while bluefin tuna eggs measure .04 -.05" in diameter.
(Liem and Scott, 1966)).
If the giant bluefin tuna were determined only to maximize the number of its own offspring that survive to maturity, a heavier
investment of energy into a smaller number of young that were better equipped to ‘escape’ the plankton should do it. Such a large
fish should hypothetically be able to equip its young to have a considerable advantage at the start over the young produced by
smaller species. Yet it does not, nor do many other large oceanic fish. The mysterious reasoning behind this, that scientists
have not yet been able to explain, may be that reproduction in fish is to a large extent in fact designed to replenish the zooplankton
upon which they all ultimately depend.
Excerpted from a recent marine ecology text is this passage regarding benthic invertebrates, though it is equally applicable
to fish:
"We find ourselves in the somewhat awkward position of having several different hypotheses for the existence and perpetuation
of the different larval types but with the real data not seeming to fit any model completely…This must strongly suggest that
other factors are also at work in determining larval development type. Why should this be? We can only suggest that for each
species there is a necessity to balance all the factors, including energy allocation, dispersal, relative abundance of microhabitat
space, longevity of adults, and undoubtedly others, against the need for larger numbers of young. The type of reproduction that
has evolved is that which balances all of these factors with survival." (Nybakken, 1997, p 25)
"The type of reproduction that has evolved" in the ocean is, to a very great extent, one that also provides significant quantities
of high quality food, in the form of fish eggs and larvae, to a multitude of carnivorous planktivores. The small size of fish
eggs ensures that small, yet vital, consumers such as zooplankton can take advantage of them. This suggests that the "reproductive"
strategy of fish is in fact designed for this precise purpose, and that "ocean fertilization" is also an important ecological
goal of their behavior. The design is such that a few offspring will survive, enough the repopulate the parent species, but that
the vast majority will be channelled instead into the maintenance of the carnivorous web…and eggs must be kept very small, even
in the largest fish, because it is important to provide food that is appropriately sized to nourish those zooplankton that produce
herbivorous young, the tiny phytoplankton consumers who represent the web’s vital link to absorbing the essential products of
photosynthesis.
An ocean full of fish, each one putting all of its energy into the production of a small number of large, predator-resistant
offspring…intuitively this seems unsustainable, especially considering that smaller prey will be required by these fish…therefore
ultimately, maintenance of a healthy abundance of zooplankton is a basic necessity and priority for all of them. And this priority
is clearly reflected in the dominant style of their "reproductive" behavior, the production of extremely high numbers of very
tiny eggs.
THE FISH EGG: CLEVER ‘OCEAN FERTILIZATION DEVICE?’
Effective biological fertilization of the sea involves not only making nutrients available in surface waters to stimulate phytoplankton
growth, but also the simultaneous nourishing of a variety of carnivorous consumers, since their sustenance is essential to the
repopulation of their juveniles, the essential herbivores. This is what the fish egg appears to have been designed to do.
When examined from the point of view of its utility in "ocean fertilization," the fish egg appears to be a particularly clever
device. First of all, most fish eggs float. This physical movement shifts protein dense material upwards through the water column,
acting to partly compensate for the surface-depleting effect of sinking organic material. A biological "upwelling" therefore
also occurs.
Once they are positioned within the upper layers of the water, a high proportion of fish eggs are eaten before they hatch. The
fish egg is virtually defenceless against its predators, and due to its high protein and energy content, probably represents
the most nutritionally valuable food item available in the plankton. The eggs that are not consumed will, after a short while,
hatch and transform themselves into tiny fish larvae. During the time (days to weeks to months) that it must live in the plankton,
the larva feeds on edible matter that is smaller than itself, accumulating nutrients and energy within its body while slowly
and continuously releasing an appropriate dosage of fertilizer for the phytoplankton. As it grows, the fish larva presents itself
as a feeding option to a range of ever larger carnivores. Sustaining these carnivores allows those species to continue to live
and reproduce, to cycle nutrients back to the phytoplankton, and to feed their own consumers.
So the fish egg can be appreciated as a cleverly designed and important biological ocean fertilization device. If it is not
consumed in its original presentation, it will automatically transform itself into a slightly different food item that is suitable
for an ever-increasing size range of predator. In effect, then, the fish egg can be seen to provide fertilizer not only to phytoplankton
and zooplankton, etc. (and most of the food value is extracted at these lower levels), but also a rippling fertilizing effect
that is ultimately felt throughout the entire web.
CONCLUSIONS
1. The decline in zooplankton production can plausibly be related to the near total removal of many spawn producing marine
species from the ocean, an end result of centuries of human fishing. Early failure to recognize the highly carnivorous nature
of the zooplankton has perhaps hampered scientific understanding of the significance of ‘animal’ food to zooplankton, including
the floating spawn produced by larger organisms.
2. The zooplankton assemblage functions as a "keystone" in the marine ecosystem. It is the link between photosynthesis and the
carnivorous web, but its success depends on the continuance of both sides, not just the phytoplankton. Disabling the zooplankton
is equivalent to punching out the "keystone" from an arch; its removal will guarantee the collapse of the entire structure. The
ultimate result of disappearing zooplankton will be the disappearance of many marine species beyond those that we have specifically
targeted by fisheries.
3. Physically forced phytoplankton blooms will predictably persist in an ocean with depleted zooplankton stocks, but efficient
uptake of nutrients, fixed carbon and energy into the living web will not. Relatively more sinking, decomposition and denitrification
of new organic material will occur instead. Measurements of seawater "greeness" or chlorophyll concentrations can provide no
reassurance that primary production is occurring at particularly high levels.
In ocean areas and seasons where surface waters tend to become nutrient depleted, this tendency will increase (out of proportion
to the increase predicted by rising temperature alone). Species with lower nutrient requirements will become relatively more
dominant in the plankton of these areas. Blooms of toxic cyanobacteria and species associated with them (some dinoflagellates)
will increasingly form in zooplankton-depleted surface waters, often during summertime. The recent global rise in harmful algae
blooms and the occurrence of toxins in seafood is therefore predictable.
4. Protection and maintenance of the zooplankton is vital. Recognizing the extent of their carnivorous nature, and their vulnerability
to food limitation, should help us to realize that maximizing the production of ‘all spawn’ in the sea needs to become a management
priority. By this reasoning, all continued fishing on spawn-producing species puts the entire ecosystem at increasing risk (i.e.
removing large quantities of crustaceans, such as lobster, crab and shrimp, from the fish-depleted and zooplankton-depleted North
Atlantic ocean is probably very dangerous). Removing these crustaceans, even though they are currently relatively abundant, invites
total collapse of the ecosystem. Their continued existence, and their contributions of spawn to the zooplankton, are probably
helping to prop up the faltering zooplankton population.
From the point of view of the zooplankton, one egg or larva is essentially just as good as another (all will be happily consumed
- size, not species, determines the acceptability of these food items to predators). Therefore, greater numbers of crustacean
spawn may currently be providing partial compensation for the missing numbers of fish spawn. This needs to be recognized and
considered in planning "ecosystem management." Fishery managers are struggling, without much success, to move away from a "single
species" assessment and management style to a comprehensive "ecosystem approach." It is time for a radical leap in thinking,
and ‘zooplankton conservation and protection’ should provide a useful concept.
If the zooplankton decline continues, it is predictable that a threshold will be reached after which rapid collapse of the
system will occur. An adequate food density is critical to the survival of all of the organisms involved, and the ‘watering down’
of the zooplankton will not be tolerated beyond a critical level. Blaming this trend entirely on "global warming" is unfortunately
reminiscent of the excuses that we have always made for the disappearance of marine species over the years. Blaming the damage
on "global warming" is another way for fishery managers to say "there’s nothing that we can do about it." FISHING is now clearly
implicated in this global disaster. It’s time to face up to the reality of the extent to which this activity of ours has contributed
to the utter ruin of the ocean.
5. Recognition of "tight cycling of nutrients" as a principle that operates throughout the entire marine ecosystem, not the
just tropical areas, allows us to begin to see all of the ramifications of centuries of fishing removals. One plausible effect
has been a long term, steady lowering of the capacity of the ocean for primary production/carbon fixation. Our relentless removal
of the spawn producing marine species has gradually undercut the supply of this carnivorous food to the zooplankton. This lowering
has taken place over centuries, and the downward trend in marine primary productivity over that time will predictably have upset
the previous ocean-atmosphere carbon balance, to the extent that the ocean has, in recent centuries, been transformed into a
carbon source. (In fact, the rising trend in global atmospheric CO2 levels matches the timing of the expansion of the commercial
fishing industry much better than it matches our patterns of fossil fuel combustion…for more on this hypothesis, see
"Strangelove
Ocean.")
6. It will be far more difficult to put the marine ecosystem back together than it was to take it apart. For several years I
have been suggesting that, besides stopping fishing, we should be ‘feeding the fish’ to aid the recovery of the stocks. Recognizing
that pouring liquid sewage into harbours and estuaries does not effectively stimulate the growth of fish stocks, I reasoned that
distributing sold edibles offshore (my best suggestion was that we somehow use human food waste) might be a better way to ‘feed’
what appears to be a starving ecosystem. That plan, if carried out thoughtfully, might possibly help to increase standing stocks
of some spawn producers…but whatever we do in our last ditch attempts at ‘ocean fertilization’ will never match the elegance
of the original design…the fish egg.
Copyright © Debbie MacKenzie, 2002
SIGN GUESTBOOK...enter your comments or read others' comments... VIEW GUESTBOOK REFERENCES Bertness, Mark, D., Steven D. Gaines and Mark E. Hay (eds), 2001. Marine Community Ecology. Sunderland, Massachusetts: Sinauer Associates Inc., Publishers. DFO, 2000. State of phytoplankton, zooplankton and krill on the Scotian Shelf in 1998. DFO Science Stock Status Report G3-02 (2000) Hempel, Gotthilf. 1979. Early Life History of Marine Fish: The Egg Stage. University of Washington Press. IGBP Report 40. GLOBEC Report 9. 1997. Ocean Ecosystem Dynamics (Globec) Science Plan. Stockholm, Sweden. Kerfoot, W. Charles and Andrew Sih. 1987. Predation: Direct and Indirect Impacts on Aquatic Communities. University Press of New England. Lasker, Reuben (ed) (undated book(!), circa 1980) Marine Fish Larvae-Morphology, Ecology, and Relation to Fisheries. University of Washington Press. Liem, A. H. and W. B. Scott. 1966. Fishes of the Atlantic Coast of Canada. Fisheries Research Board of Canada, Bulletin No. 155 Messieh, S. 1988. Spawning of Atlantic Herring in the Gulf of St. Lawrence. American Fisheries Society Symposium 5:31-48 Nielson, E. Steemann. 1971. "Plankton Production" in Nybakken, James W. 1971. Readings in Marine Ecology. Harper and Row Publishers, New York. Nybakken, James W. 1997. Marine Biology: An Ecological Approach (4th edition). Addison-Wesley Educational Publishers Inc., USA Orr, A. P. and S. M. Marshall. 1969. The Fertile Sea - being the Buckland Lectures for 1957. London; Fishing News (Books) Limited. Polunin, Nicholas V. C. and Callum M. Roberts (eds). Reef Fisheries. London: Chapman & Hall, 1996 Steidinger, Karen A. and Linda M. Walker. 1984. Marine Plankton Life Cycle Strategies. CRC Press, Inc., Boca Raton, Florida Zhong, Zheng, et al. 1989. Marine Planktology. China Ocean Press, Beijing ______"Oral History...Straight From the Whale’s Mouth" (online at http://www.discovery.com/news/features/whales/whales.html
)
|