Fisheries models fail
to grapple with reality
(…if in doubt, should we change the reality?)
Scientists at the Canadian Department of Fisheries and Oceans have recently discounted evidence in their longest-running plankton data set (which showed zooplankton declining as fish stocks failed) because the observed trend was in stark contrast to the predictions of a favoured ecosystem model. Rather than discarding or changing the model, DFO discarded their plankton data instead, and scientists have accepted the model’s prediction that zooplankton must have increased as fish stocks failed. Drifting farther from reality, this exercise is getting us nowhere, and the model offers no insight into the cause of the continued depression of the fish stocks. I visited DFO recently, and tried to suggest a way to build a new model…
- Debbie MacKenzie
September 22, 2004
|
|
|
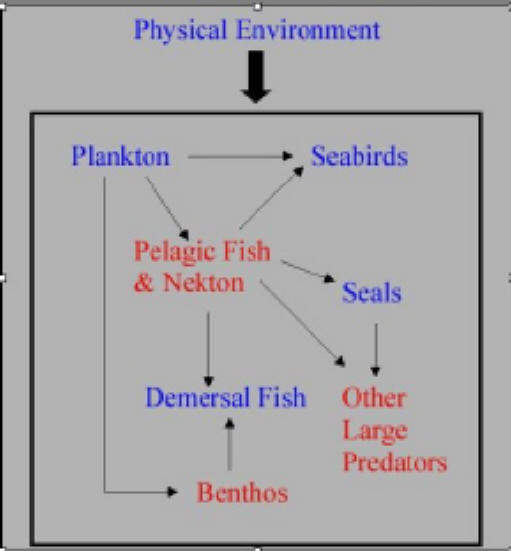
DFO model of fish production (click to
enlarge) - the rate of overall food production is determined solely by the
"physical environment" (climate). |
|
|
|
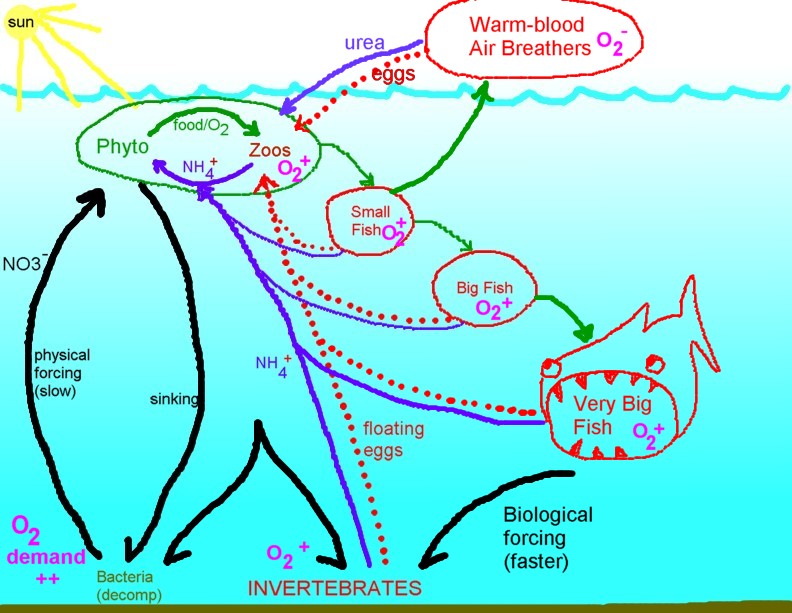
Model of fish production showing both
"physical" and "biological" routes that stimulate food (phyto) production.
These dynamics are additive. Fish are integrated into a living web that
raises the rate of overall food production to a higher level than the rate
"forced" by the physical environment alone. If this is a significant
dynamic, then fishing has some potential to slow system productivity
overall. A robust animal component also contributes to greater oxygenation
of seawater. |
|
|
|
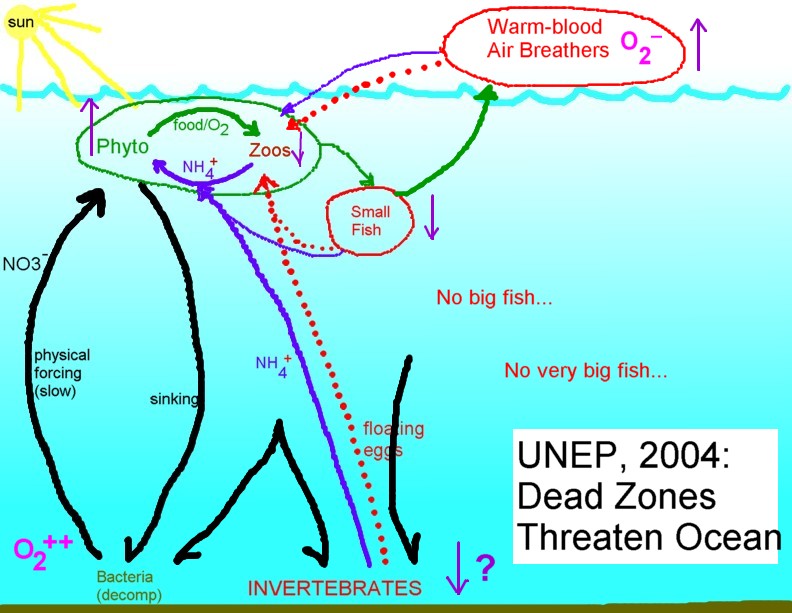
The ecosystem as it has been altered by
fishing. A "bare bones" animal web still operates, but at reduced
strength. Productivity is lower but bacterial cycling, and the associated
loss of oxygen, is more prominent. |
|
|
|
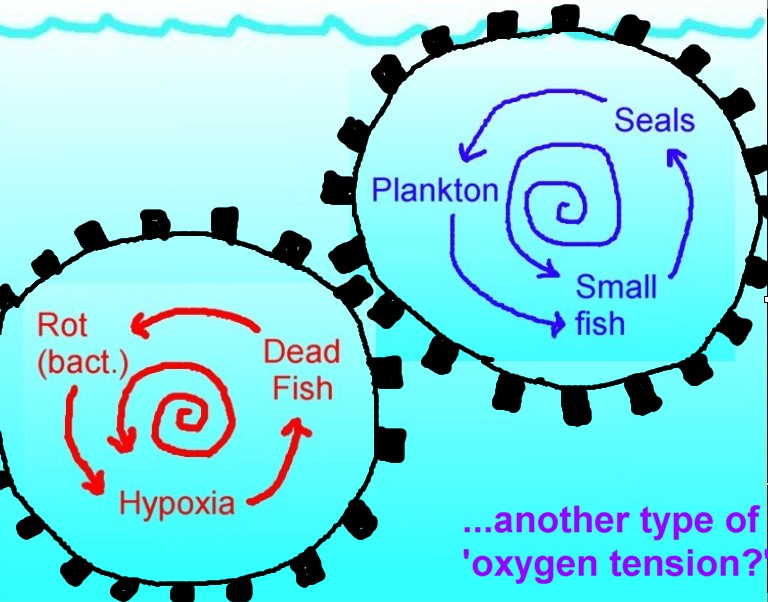
Illustration of the concept of
competing cycles, each with an inherent tendency to accelerate. Red cycle
(bacterial decomposition) can snowball into a "dead zone." Blue cycle
(interconnected animal web) also self-accelerates due to internal positive
feedback, and as a whole this tends to suppress the red cycle and to
increase the quantity of both animal life and oxygen in the ocean. Some
animals access the atmosphere, offering an oxygen-sparing marine metabolic
route in extreme contrast to the bacterial action, which heavily depletes
oxygen from the water. Seals, whales and seabirds therefore naturally
assist the "living ocean" to ward off fish-killing "dead zones." The "dead
zone" problem in the accepted scientific view is thought to be driven only
by nutrient pollution. However, removal of the animal web can also be seen
to aggravate this tendency. The red and blue cycles compete for dominance
- and cutting down the blue gives an edge to the red. Facilitating a
rebuilding of strength in the blue cycle, however (allowing marine animal
life of all types to naturally re-establish themselves and accumulate
biomass in the sea), will tend to force "dead zones" to recede. |
|
**
DFO's author, Bundy (2004), identified as a major concern the discrepancy
between the Ecopath model requirement that small zooplankton (SZP) had
increased while independent plankton estimates suggested it had decreased
instead. Such a degree of uncertainty about this crucial foodweb element
threatens to render all results of the modeling exercise irrelevant. It is
interesting to note that the diet data on types of fish that consume SZP
directly showed a declining proportion of this particular food in their
diets, when comparisons were made before and after the groundfish
collapse. (This declining trend was shown for small demersals, capelin,
sand lance, pollock, redfish, spiny dogfish, haddock, large and small
silver hake, and large and small cod (Bundy, 2004). Other data from
capelin and mackerel in the Gulf of St. Lawrence show the same trend.) A
general shift toward eating a larger proportion of fish, an "increase in
piscivory," was noted in many fish species, and it was suggested that this
trend naturally followed from the increased availability of forage fish.
But is that it? ...there are reasons to doubt
the reported dramatic increase in the biomass of forage fish, including
diet shifts within the forage fish. A fish diet shift away from SZP might
also be expected to follow from a declining availability of SZP, a food
that is more easily acquired than larger prey (which might need to be
chased down before being eaten). Some fish, like mackerel and capelin, can
eat SZP by filter feeding, and they can also eat larger foods by catching
these one piece at a time. For these fish, SZP has long been their main
food, but it has recently declined in prominence in their diet. It is hard
to imagine that SZP concentration could increase in the environment and
that these fish would choose to eat less of this virtually stationary
food, preferring to expend their energy chasing more difficult prey. It
seems to me that the increase in piscivory in such species most likely
reflects a declining availability of SZP rather than anything else. Many
fish that exhibited the increase in piscivory have also shown declining
growth rates, suggesting more difficulty feeding. The sum of changes is
simply incompatible with the conclusion that food availability for fish
has generally increased. A decline in SZP is an ominous signal of decline
in the health of a "fish-producing ecosystem" ...and DFO urgently
needs to get to the bottom of this question! |
The Honorable Geoff Regan, M.P.
Minister of Fisheries and Oceans, Canada
200 Kent Street
Ottawa, ON K1A 0E6
Min@dfo-mpo.gc.ca
Dear Mr. Regan,
On September 15th I was given an opportunity to present my alternate model of “how the ocean works” and “what is wrong with fish stocks today” to a group of DFO scientists at the Bedford Institute of Oceanography. We spent several hours discussing the issues. I also received written feedback from the scientists. While we agreed on the seriousness of recent negative changes in the marine ecosystem, and seemed to share a concern that worse developments are likely to occur should the downward trends continue, the scientists remained extremely skeptical that there could be any significance to the ecologically damaging dynamic that I was proposing (an erosion of ocean fertility by fishing).
I tried to show how the evolution of life in the sea, by the accumulation of ever greater numbers and sizes of fish and other marine animals, naturally led to increasing efficiency of marine nutrient cycling due to the normal activities and movements of the animals. Basic linear models (plankton => small fish => big fish) do not capture this effect. Zooplankton are crucial links in the process that channels energy into fish (at least we agreed on this point) and the very presence of these tiny grazers causes an acceleration of plant (phytoplankton) growth, because zooplankton “regenerate” (excrete) plant fertilizing materials. We agreed on this point too.
However, a key concept that I tried to convey is that the larger animal web as a whole has a similar, additional growth-stimulating effect because this living reservoir itself “regenerates” (excretes) zooplankton –- and the release of billions of excess floating spawn (eggs) is one key pathway by which this is accomplished. Maintaining a robust zooplankton cohort can be viewed as a co-operative strategy of all larger marine animals which provides “pay back” by maximizing the amount of energy (food) flow, from photosynthesis to fish food, that occurs in the sea. Bottom invertebrates, like starfish, clams, and worms, are the major players, excreting phenomenal numbers of tiny floating eggs, but fish, and even air-breathing fish predators such as seabirds and marine mammals, are integrated into this synergistic zooplankton-regenerating dynamic. Two main effects are: zooplankton-enhancement by animals has a knock-on phytoplankton-regenerating effect (which raises overall system productivity), and it enables more efficient capture of energy, and the diversion of this energy into the animal web, as opposed to the older bacterial breakdown cycling route. (This is how a sea full of fish is more naturally resistant to forming “dead zones,” in which oxygen is depleted by a high rate of bacterial decomposition of organic matter. This is a new interpretation of how the ocean works, that shows fish woven into a web that has some capacity to regulate its own productivity. Broad, negative ecological effects might therefore be theoretically induced by fishing, including a decline in system fertility. This concept threatens the status quo of fisheries and fisheries science.)
The main scientific objection to my model seemed to be that nutrient cycling through fish represents such a minute fraction of the total, that the removal of all fish would be expected to have no detectable effect on ocean fertility. However, the removal of all of the fish has the potential to inadvertently cause a considerable drain on the population of bottom invertebrates too, along with other important reserves, because these share a common link in zooplankton regeneration. I may have made this clear at the meeting, or perhaps I did not. It is very complex.
The biggest surprise to me at this meeting was the realization that DFO scientists are not now greatly concerned by the declining zooplankton counts that they have observed to span the last four decades, because they have decided to discount this data (from the “Continuous Plankton Recorder,” which indicated that a decline in zooplankton abundance occurred as the fish were declining.). Why does DFO now disbelieve their own data? The scientists now argue that the documented decline in zooplankton counts is unlikely to be significant because the monitoring equipment was relatively crude and inadequate. (But they have nothing else that extends back for any length of time…) Regardless, it seems to me that an important reason for the recent discounting of the Atlantic Canadian zooplankton data may relate to the impossibility of reconciling this data with currently used ecosystem models. In one such recent exercise (using “Ecopath” to model the ecosystem of the eastern Scotian Shelf before and after the groundfish collapse), it was necessary to accept that an increase in zooplankton biomass had occurred concurrent with the loss of fish in order for the model to be made to balance (Bundy, 2004)**.
The exhaustive “Ecopath” modeling exercise produced no explanation for the ongoing failed rebuilding of the groundfish stocks, and it brought DFO no closer to understanding why large cod and other fish are rapidly dying off instead. Is this because DFO scientists are still struggling to understand the ecosystem with models that lack some crucial truth about how the ocean works? I am convinced that a significant decline in zooplankton has in fact occurred for many reasons, including the starved condition of the remaining big cod, the gradual decline of plankton-feeding barnacles and other invertebrates along the coastline, and the apparent confirmation of declining zooplankton in the CPR data. The most important lesson from the Ecopath modeling exercise might be that so many elements are shifting far outside the ranges of where they are “supposed” to be, that this model fails to capture what is really occurring. Rather than blaming the gross misfit on deficient data, serious deficiencies in the model itself should now be suspected. Instead of adjusting the zooplankton counts to fit the model, perhaps the model ought to be adjusted to accept the actual zooplankton counts.
At the meeting I tried to explain, albeit briefly, how observations of long-term changes that have occurred along the shoreline can help scientists trying to diagnose the changing condition of the wider coastal ocean ecosystem (unexpected declines in barnacles and other invertebrates, signals of declining fertility in seaweeds). No scientific monitoring of these changes has been done, and the DFO scientists made virtually no comment on this aspect of the picture, or its possible significance. The shoreline observations that I offered fall far outside the expectations of the accepted model. Systematic scientific investigation of changing trends in near-shore marine organisms should become an important priority for DFO science, because many of these organisms offer unique advantages as indicators of overall marine fertility and of plankton productivity, and because these species have shifted in such dramatic opposition to the expectations of the accepted models that this knowledge should have useful extensions in improving the scientific understanding of “the big picture.”
I was told that shoreline monitoring has not been done because of a lack of funding, and that the science program itself is in danger of being scaled back further.
(Mr. Regan, the DFO science program badly needs to be re-organized and re-focused. Monitoring critical ecosystem health indicators is what most needs to be done, yet DFO is still not sure what these are (beyond climate variables). Certain recent initiatives, such as the $6 million devoted to studying seal diets, are a waste of resources. Seals eat fish, but they cause no harm to the ocean that produces fish. In fact, they are naturally beneficial. If we know this much, then we do not need to dissect and scrutinize seals to determine the precise mix of fish species they are eating. On the other hand, tracking indices of ocean fertility, with a close focus on all types of algae, and on oxygenation and zooplankton, will provide relevant information on trends in the capacity of the ocean to produce fish and to provide other “ecological services” to the planet.)
The meeting a BIO was valuable to me because it confirmed my suspicion that the DFO scientists would offer no objections other than ones I had already heard. My impression was that we ended up more or less where we started: the scientists are unconvinced that my “alternate model” of what drives organic growth in the sea can add anything useful to what they already know and are trying to work with, and I remain unconvinced that the tools these scientists are working with are up to the job of solving the puzzle because they simply cannot account for so many broad, significant changes, and because they have failed to incorporate all of the natural ecosystem dynamics involving marine animals.
I emphasized how alarmed I am by the ongoing negative changes in ocean life, how I am convinced that fishing has been the major cause of the broad ecological damage, and how extending moratoriums on fishing is what now must be done if we hope to stop the destruction or to ultimately salvage any of the larger marine animals. None of the scientists openly disagreed with these sentiments, but it was made very clear that the freedom to voice such opinions is not a part of working for DFO’s Science branch. (I said: “Aren’t you extremely concerned about where all of this is heading? When are you going to sound the alarm, to push the emergency buttons, and bring a halt to the ongoing destruction being caused by the fisheries?” The reply: “We are not allowed to say that!”) Scientists do not set the policies, and their conversation with me was not supposed to stray into that contentious territory. This much was made very clear, both during and after the meeting. That is too bad.
It seems to me that the compulsory priority of supporting commercial fisheries unavoidably distorts the work of DFO science. When the sticky question that needs answering (how has fishing upset the workings of the entire ocean ecosystem?) threatens the future of the commercial fishing industry, are DFO scientists allowed to freely pursue the question? The answers you find will inevitably be shaped by the questions you ask. (Whereas an unemployed person like myself can ask any question, and can freely stray from scientific details to policy issues, and back again, if it suits me…)
Here is part of what I see: pro-fishing interests are still in the driver’s seat, still controlling DFO policy, and they are currently taking advantage of the “well, we just don’t know enough yet” messages emanating from the Science branch. As long as science “doesn’t know,” then the default setting for fisheries management is to carry on.
Two things that are legally supposed to be occurring now in marine management in Canada, “erring on the side of caution,” and the “reversal of the burden of proof,” are not being done – NOT AT ALL. If in doubt about possible adverse ecological effects, fisheries are not supposed to be prosecuted (and yet, we have nothing but such doubt). The “burden of proof” is not supposed to be on those who would protect marine life, but rather placed on those who would exploit it, to show that further exploitation will not harm either the resource itself or the system that produces it. This means that the onus should now be on the fishing industry to provide proof that their activities are causing no harm. But they cannot do that, and practically, this requirement is ridiculous. All the pro-fishing lobby can point to is the current high abundance of certain commercial species, such as lobster, shrimp and crab…but this gives us no indication whatsoever of the “ecosystem” effects of removing significant numbers of them. Hanging on to the “single species” approach is the only strategy that can be used to justify continuing with commercial fisheries, even though this approach has been widely recognized as inadequate and inappropriate. (Remember how spectacularly it failed with cod?) But a genuine shift to “ecosystem management” will shatter current fisheries, if it is done honestly. So, what is DFO to do?
Bend the numbers to fit the ecosystem model that provides a rationale for thinking that fisheries are basically “sustainable?” (…in a test of whether or not we can successfully force nature into a “box” we have created that allows us see what we want to see? This is folly.)
Mr. Regan, DFO Science is not getting any closer to solving the case of the missing fish. My impression is that the scientists are in a rut, and they are spinning their wheels. But, certain “powers that be” in DFO seem content to leave them there, because the economic goals of commercial fisheries can continue to be pursued in the meantime. These activities are justified because DFO science officially “doesn’t know” what damage fishing might cause to the ocean. Scientists “don’t know” because they are not asking the right questions – one specific example of this can be seen in the failure of DFO Science to investigate the extent of ecological damage to bottom life caused by bottom trawls. The larger ecological question that I have raised (“what degree of damage has fishing caused to the very foundation of ocean life, the plankton?”), seems to stand no chance of being addressed in this environment.
DFO and I are nowhere near to being on the same page.
There is a time for slow, methodical study, but there is also a time for decisive action. This situation reminds me of a team of doctors puzzling over a sick man. Interesting changes in the patient’s blood are being studied microscopically and through a battery of biochemical tests. The precise pathology is unclear. However, some key health indicators are noted to be shifting dangerously. The oxygen content of the man’s blood is falling, his blood pressure is declining too, and the medical team realizes that their patient faces imminent, irreversible “shock,” and likely death, unless appropriate corrective measures are taken immediately.
Continuing to remove blood samples in a methodical search for the exact identity of the responsible virus or toxin, for instance, will not save this man. This sort of investigation is worthwhile, of course, but in an acutely threatening situation, when life signs become unstable, more basic protective manoeuvres become the first priority. If the patient cannot maintain blood pressure and oxygen levels, then prompt action to counteract these problems becomes imperative. The doctors’ gross understanding of how the patient’s “system” works leads them to suspect blood loss as a possible cause of his general deterioration, and they correct the problem with a blood transfusion. Such caretakers will not allow substantial quantities of blood to continue to be removed from their weak, unstable patient without replacement. Of course, another possibility is that the caretakers will fail to realize the significance of the changing vital signs, and persist in studying the problem only through a microscope, only to be baffled in the end by the “sudden” demise of their patient.
The methodical, ongoing studies being conducted by DFO scientists are proceeding too slowly to save their “patient,” the living ocean. It is nobody’s fault, but this branch of science lacks a confident knowledge of the gross functioning of the living ocean system. If they possessed this, scientists could react with sound “ecosystem” advice in a scenario with dwindling general health indicators, such as declining ocean oxygen content (a trend that is widely evident today.) Falling seawater oxygen from any cause, as an example, should trigger an increase in the level of conservation of all marine animals…but…this would need to be based on a prior understanding of the net positive effect of animals on the oxygenation of seawater. This aspect of the ecological role of marine animals has not been included in the accepted gross models of how ocean life works, and such decision-making is about as far from the classic “single species” management approach as we could go…so, marine animal conservation decisions are most unlikely to be made on such a basis in the near future. However, I am convinced that this is the direction in which the “ecosystem-based management of living resources” needs to move.
Unfortunately, my sense of urgency about all of these things seems to be considerably greater than that of the scientists. They suppose that I am unduly worried about the effects of fishing because I “don’t understand enough,” while I think it is because they “don’t understand enough.” We are at loggerheads (…but the real loggerheads, the turtles, and other large marine species are the ones quietly facing extinction). In Canada, DFO’s ocean experts are evidently “not allowed” to make observations like mine: that fishing has already caused immense damage to ocean life, and that it should be drastically curtailed now as a legally sanctioned “precaution” against causing greater damage. Such a law does exist. But until ocean science in Canada emerges from DFO’s rut, we can only expect to see seals and fish continuing to be counted and killed in the usual manner, without any consideration of the effects that their removal might have on failing fundamental ecosystem health indicators such as oxygen content or plankton indices. Oxygen production in the ocean is related to plant fertility, and both the causes and the manifestations of ocean plant fertility are very poorly understood by marine scientists today. Some might be inclined to disagree with me, but chances are that they have not thoroughly examined the seaweed. However, in this century, we should be aware that if the living ocean ecosystem edges into a condition analogous to physiological “shock,” then our mysteriously ill patient might “die” quite suddenly.
Mr Regan, I urge you again to consider a separation of the mandates of “fisheries” and “oceans.” Canada needs “oceans” scientists that can make their first priority “the health of oceans,” and not necessarily “the health of fisheries.” A healthy ocean is infinitely more valuable than the short-term commercial gains that can still be made from fisheries. One problem is that the public just doesn’t “get it” yet that overall ocean health is crumbling, nor do they “get it” how far-reaching the effects of this will be. The future will not be a repetition of the past, and the romantic Canadian coastal “tradition” of fishing is already history. The future for many marine species, and for the people who still have their hopes pinned on commercial fisheries, promises to be surprisingly bleak…but perhaps we can pretend otherwise for some time yet, for as long as government scientists officially “don’t know” what is happening to the fish and to the ocean.
Mr. Regan, thank you again for arranging the meeting for me at BIO. It was helpful. If possible, I would like to meet with you again when you are next in the Halifax area.
Sincerely,
Debbie MacKenzie
Reference:
Bundy, A. 2004. Mass balance models of the eastern Scotian Shelf before and after the cod collapse and other ecosystem changes. Can. Tech. Rep. Fish. Aquat. Sci. 2520: xii + 193 p.
Feedback received from the scientists at our meeting (with my comments, in italics):
Date: Fri, 17 Sep
2004 14:05:31 -0300
From: "Harrison, Glen" <HarrisonG@mar.dfo-mpo.gc.ca>
Subject: DFO feedback to Starving Oceans meeting
To: 'Debbie MacKenzie' <debimack@auracom.com>
Debbie,
I want to respond to the point you made to me after our discussion on Wednesday - that you didn't think we would give your new model any consideration. I don't want you to be left with the impression that we are unreceptive to new ideas and ways of thinking about things - we are constantly testing new (and existing) theories. However, to be absolutely honest with you, I don't think DFO will adopt your model but there are scientific reasons, many of which we highlighted at the meeting. There was so much discussed that the major points might have been lost. Therefore, I'll try to summarize and maybe you can think about these as you fine-tune your thinking on this complex problem.
First of all, I want to emphasize that your observations on the changes in our marine ecosystems and the questions you raise are all valid - we have absolutely no issue there. The problem comes in with your efforts to tie these all together through a single process - ocean nutrient cycles, i.e. the "starving" ocean concept.
Your argument is based on the premise that the removal of large fish has disrupted the natural "biological" flow of nutrients that are required for primary production with the consequence that the system cannot produce enough energy to sustain itself. You go on to describe where the biological nutrients come from (or rather came from since the system has lost its big fish) and how that has fed the system. You wrongly dismiss physical sources of nutrients. (No. I said that the biological nutrient supply augments the physical supply.) The nutrients you identify are biologically produced/recycled forms (ammonia, urea, dissolved organic matter). You invoke a couple of mechanisms for getting those nutrients to the surface where phytoplankton can use them: (1) zooplankton migration and (2) turbulent mixing by rising fish eggs. Here's where we start having problems with that model. In order for this to work, we need a reservoir of ammonium, urea and dissolved organic matter down where the big fish are(were). (No you don’t, not really, because the big fish themselves “were” the reservoir. Think of those fish plus all of the bottom invertebrates as ammonium and zooplankton slow-release devices.) When we look at the distribution of these nutrients, however, there is no reservoir of any of those at depth, i.e. ammonia nor urea accumulate in seawater (except in hyper-eutrophic coastal waters) and dissolved organic matter reservoirs in the ocean are, in fact, at the surface and concentrations decrease with depth. (Actually, DOM concentration does not decline nearly as dramatically with depth as does POM, although a decreasing tendency is evident. I have found this documented elsewhere. But do you have any data on DOM distribution in Canadian waters? I have requested this information from DFO repeatedly, but have never received an answer.) The only source that can be tapped at depth is nitrate (and I don't think you'll find any literature that says that zooplankton absorb nitrate). (Neither do I. Actually, I have taken more than a passing glance at this question, and it seems to me that enough unknowns remain in zooplankton ecology to allow for the possibility that they do access DOM at depth. See my article: Vertical Migration of Zooplankton: a biphasic feeding strategy that enhances new production?)
And while we are talking about zooplankton, the published data on zooplankton nutrient transport (and there are papers published on this) show that they are net exporters of nutrients from the surface ocean to depth (as Erica said), not importers. Erica and Doug have put together a more detailed discussion of zooplankton and I'll send that to you in another email. As far as fish eggs rising and turbulent mixing, I understand that some speculation on the ecological significance of fish eggs has been considered by oceanographers (e.g. Rothschild, Bioscience, Vol 31, 1981) but the nutrient transport angle has not been pursued.
More fundamental to your model, however, is the point I tried to make a number of times at our meeting. We are not questioning whether many of the pathways of nutrient supply you described exist, we are questioning, "how important are they"? Nutrient cycling probably has been altered by changes in the ecosystem in our waters but have those changes affected primary production? You talked about the trophic pyramid and how each feeding level contributes to nutrient regeneration to fuel phytoplankton growth. I'll use that to make my point. Let's take a very simple case where there are 1,000 nitrogen-units of phytoplankton on the Scotian Shelf. Based on well-established tropho-dynamic theory, the phytoplankton would produce 100 N-units of Calanus that would produce 10 N-units of capelin that would produce 1 N-unit of cod. Remember, it takes 1,000 units of nitrogen to sustain the phytoplankton. Now, let's remove ALL the cod - thus you have lost the systems capacity to deliver only 1/1000 of the nitrogen required to sustain phytoplankton growth. (Good grief Glen! - did you say "let's remove ALL the cod?!" Gads! - there's the problem!...sorry, I am actually taking this very seriously... ) There MUST, therefore, be an external source of nitrogen to keep the phytoplankton growing and I would argue that nitrate delivered from horizontal and vertical mixing by physical processes is that principal source. (But your nitrate-driven model clearly misses something, because between the 1960’s and the present it looks as if there has been a functional shift from “1000 N-units of phytoplankton producing 100 N-units of Calanus” to something like “3000 N-units of phytoplankton producing 50 N-units of Calanus” – according to CPR counts – I have still seen no explanation for the divergence between phyto counts and zoo counts…except for what I heard at our meeting: that your scientists no longer accept that your falling zooplankton counts represent a real decline in zooplankton biomass.) Don't misunderstand me, recycled nutrients are important for phytoplankton and probably fuel as much as half of the primary production on the Scotian Shelf (mostly in summer). What we are saying is that big fish are not the important nutrient source (major contributors are microbes, then zooplankton, then forage species in that order of importance). There are simply not enough of the big fish to change the balance of regenerated (biological) versus new (physical) nutrient supply. (Part of what I am saying is that big fish are integrated into a pattern whereby large animal biomass replenishes zooplankton, and therefore contributes to optimizing primary production, which the zooplankton do. And there were once MANY more big fish…before someone REMOVED ALL THE COD!)
We are not arguing that big fish are unimportant for the integrity, energy flow, stability of the ecosystem, we're simply saying that you can't make the case they are important as sources of nutrients to sustain primary production. The same argument would hold for the oxygen and carbon dioxide cycles that you mentioned in your presentation. Physical processes and metabolic activity of the small critters (microbes, zooplankton) are the major players here. In a nutrient context, therefore, our view is that the oceans off our coast are not starving despite the dramatic changes that have occurred in the ecosystem (reduction in large fish). (Again, big critters do not only eat them, but they also affect the metabolic activity of the smaller critters, because in effect they “regenerate” zooplankton – echoing the effect that the little critters exert on the smaller plant forms that they eat – therefore big fish are closely linked to the “major players,” making them perhaps “important” players themselves?)
I'd like to leave you with a couple of more general points that Charles Hannah and Alain Vezina raised during our discussion. DFO scientists are working cooperatively with scientists worldwide on these complex problems and taking leadership roles, e.g. in trying to understand the linkages between environment/climate changes and marine ecosystems, in developing a framework for an ecosystem approach to resource management, etc. (This is fine, but I do not see a similar level of attention being paid to studying the linkages between FISHING and marine ecosystems. Changes in marine life are simply not all about “environment/climate changes,” and this can be seen particularly clearly in Atlantic Canada. Instead of following the international pattern, why not start something in a new direction? I can't help wondering if at some level political forces are helping to keep the science so focused on "environment/climate" instead of FISHING. Note: this comment is not the same as dismissing the significance of environment/climate.) The reason we are important "players" is that we conform to a well-established, rigorous scientific process that carefully weighs all lines of evidence before coming to a conclusion about cause-effect. We entertain new ideas but they have to be subjected to this rigorous process. Your theory, although plausible on the surface, is simply not defensible when all factors are considered. Keep up the good fight - just don't rely on nutrient starvation as your scientific underpinning. (Look, the cod are still starving and you have not provided a reason why! But I have. Please consider that your “scientific underpinning,” your classic climate-forced production model, is not as solid as you assume it to be. Has your “rigorous process” not caused you to doubt this yet?)
I have consulted all the DFO scientists present at our meeting before sending this to you so you are certainly welcome to include this on your website if you want to summarize the feedback you received from DFO scientists. (Thank you. It was nice to meet with you, but honestly, I felt to quite an extent that I was only being “humoured” in being allowed to speak with your scientists. Your inappropriate criticism that I “wrongly dismissed physical sources of nutrients” suggests to me that you were not really listening to what I was saying. Your mind may have been made up before I arrived. Final thoughts: I strongly urge you to look at what is happening along the shoreline. ...and "The problems we have today will not be solved by thinking the way we thought when we created them" (Einstein) - cheers, Debbie)
Regards,
W.G.(Glen) Harrison,
Head
Biological Oceanography Section
Ocean Sciences Division
Bedford Institute of Oceanography
Box 1006, Dartmouth, Nova Scotia
Canada B2Y 4A2
Ph: (902) 426-3879
Fax: (902) 426-9388
Email: harrisong@mar.dfo-mpo.gc.ca
A few more comments were received on zooplankton:
Zooplankton in Debbie's model
1) Debbie suggests that zooplankton that perform vertical migrations become infused with nutrients at depth, due to hydrostatic pressure, and bring them back to the near surface layers to fuel phytoplankton growth.
The first thing to recognise is that there are two classes of zooplankton: microzooplankton (<200um in size, 1mm = 1000um) and mesozooplankton (200-2000um in size). Both feed and excrete ammonia in the near-surface layers. Microzooplankton regenerate much more ammonia than mesozooplankton, and they do not migrate to depths where nutrient (nitrate) levels are high.
The mesozooplankton is dominated by copepods (70-80% of the biomass) and many of these do undertake daily vertical migrations, but for the dominant spring species (Calanus finmarchicus) and for most of the small species, which dominate in summer, these migrations are not extensive (ca. 10-20 m or less). A few copepod species and euphausiids migrate to depths of >100m, and because of this they are only found in the shelf basins. (This is an interesting fact, no? What advantage does the depth of the water column offer to these species? They do experience a greater pressure gradient.) Most copepod species feed most actively at night, when they are at their shallowest depth, and they excrete ammonia continuously, more rapidly when they are feeding. (This too, supports my hypothesis that ammonia produced by metabolic activity at depth is disproportionately excreted in the surface water – therefore ammonia is retained for a while in the bladders, and conveniently it offers a bit of positive buoyancy. Filling the gut will increase internal pressure and cause the bladders to empty during feeding – this seems more likely than that ammonia generation is faster during feeding. See my article on this topic: Vertical Migration of Zooplankton: a biphasic feeding strategy that enhances new production? ) They cannot allow ammonia to build up in their body tissues, because it is toxic. The vertical migrators tend to have long gut turnover rates, so that they still have food in their guts for part of the day after they descend, and overall they provide a net downward flux of particulate (in faeces) and dissolved (ammonia and dissolved organic material (DOM)).
I now understand that when Debbie said that "all crustaceans have bladders" she was not referring to organs such as the swim bladders of fish, although she was implying that they had something to do with buoyancy. Instead, I think that she meant was talking about organs corresponding to bladders in humans, i.e. excretory organs. Assuming this, then she is right, they are located somewhere in the head region. There is no reason to suppose that these organs would ever reverse their role and take up liquid, however; they are not suited to do so. Also, there is no reason to expect that increased hydrostatic pressure should cause infusion of nutrients or DOM when the migrators are at depth. These organisms are contained within an impermeable shell and they regulate what they take in, just like fish, whales and seals. There is no evidence that copepods feed on DOM, while there is evidence to suggest that they do not.
2) When eggs from groundfish float up to the surface layers, zooplankton feed on them, regenerating nutrients to fuel phytoplankton growth.
Cod eggs are ca. 1.3-1.5mm in diameter. The largest stage of the largest copepod that is found over the banks during the groundfish spawning period is 3mm in length (adult C. finmarchicus). Most copepods there are much smaller (<1mm). There is simply no way that these copepods could eat particles as large as cod eggs. In fact, there is a relationship that has been published that shows that over a large size range of copepods, they can only take particles of up to 1/18 of their length. For C. finmarchicus females this would be ca. 0.17mm, which corresponds to the size of a fairly large diatom. Large krill (3cm long) might be able to eat fish eggs, but they do not live on the banks, but in the basins, since they need water depths of >100m for their daily vertical migrations. On the other hand pelagic fish, such as herring and mackerel, are known to be voracious predators of fish eggs and larvae. (Cod eggs enrich zooplankton because they “are” zooplankton, and as living marine animals they excrete ammonia. They participate in the zooplankton picture for probably most of their short lives. Invertebrate eggs tend to be smaller and are probably more likely to be consumed directly by other small zooplankton like copepods.)
3) The abundance of zooplankton on the eastern Scotian Shelf has decreased on since the 1960s.
Debbie has suggested this based on data presented in a couple of DFO technical reports. The data come from the Continuous Plankton Recorder (CPR). This device is towed at a depth of 7 m, from commercial ships on a monthly basis. Debbie only showed the data for one species of copepod and "euphausiids". Granted, the copepod C. finmarchicus, is a very important copepod for larval and juvenile cod, as it is for the small pelagic fish, which eat it throughout their lives. Euphausiids, which are dominated by one species, Meganytiphanes norvegica, have also decreased in abundance. Another copepod species, Calanus hyperboreus, however, has shown a marked increase in abundance - and this data were presented in one of the DFO reports. (As I asked at the meeting, what is the trend in the nutritional value of the plankton, per unit volume of seawater? Many things seem to indicate to me that this is declining.)
Numbers of C. hyperboreus are low compared with those for C. finmarchicus, but this species is about ten times bigger per individual in terms of biomass, and it may not be as well represented in tows at 7 m, because it prefers relatively cold water and lives deeper in the water column. The interpretation of these data is not straightforward, but it is seems most likely that the changes are associated with changes in water masses. C. finmarchicus and M. norvegica are North Atlantic temperate species; C. hyperboreus is an Arctic species. On the eastern Scotian Shelf there has been more colder fresher water, derived from the Arctic, in the early 1990s than there was in the 1960s. Thus, it is likely that conditions on the eastern Scotian Shelf are now better for the Arctic species and worse for the temperate ones. This is not the only, or the complete, interpretation, however. (The cold water episode that occurred in the early 1990’s has been resolved now for over five years, has it not? Would you expect such a sustained effect years after temperatures had returned to “normal?”)
Another point we did not get around to discussing:
Debbie suggested that blooms of "tropical" coccoliths are now occurring in the North Atlantic and Bering Sea, because of depleted nutrient supplies.
The coccolith species that we find in the North Atlantic, the Bering Sea and the Barents Sea are not tropical species. There are some tropical species and they occur in the tropics, but there are also some "cosmopolitan" species that are quite ubiquitous in their distribution. These are the ones that we find. Also, coccolith blooms do not occur when nutrients levels are especially low. One thing they do seem to require is a shallow mixed layer, but other than that the conditions for coccolith proliferation are not well defined.
This is all I can think of, for now.
Erica/Doug
Sign My Guestbook
![]() View My Guestbook
View My Guestbook
![]()